КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
История зоологии 3 страница
|
|
|
|
От переднего полюса тела берут начало жгутики (1, 2, 4, 8 и более — до нескольких тысяч). Если их много, они могут покрывать все тело простейшего (например, в отрядах Hypermastigina, Opalinina — с. 51), напоминая тем самым инфузорий. Длина жгутиков варьирует в широких пределах — от немногих до нескольких десятков микрометров. Если жгутов два, то нередко лишь один выполняет локомоторную функцию, тогда как второй неподвижно тянется вдоль тела и выполняет функцию руля. У некоторых жгутиконосцев (роды Trichomonas,Tripanosoma, см. ниже) жгутик проходит вдоль тела (рис. 19) и соединяется с последним при помощи тонкой цитоплазматической перепонки. Таким образом, формируется ундулирующая мембрана, которая своими волнообразными колебаниями вызывает поступательное движение простейшего.
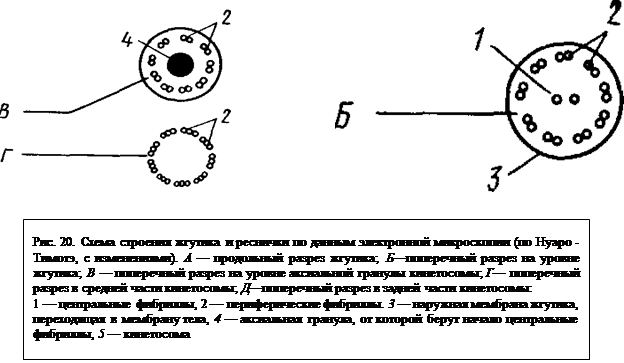
В деталях механизм работы жгутиков различен, но в основе это винтообразное движение. Простейшее как бы «ввинчивается» в окружающую среду. Жгутик совершает от 10 до 40 об./с. Ультраструктура жгутиков, как показывает электронная микроскопия, очень сложна и обнаруживает поразительное постоянство во всем животном и растительном мире. Все жгутики и реснички животных и растений построены по единому плану (за единичными отклонениями) (табл. I, 3, 4). Каждый жгутик слагается из двух отделов. Большую часть его составляет свободный участок, отходящий от поверхности клетки наружу и являющийся собственно локомоторным. Второй отдел жгутика — базальное тело (или кинетосома) — меньшая по размерам часть, погруженная в толщу эктоплазмы. Снаружи жгутик покрыт трехслойной мембраной, представляющей непосредственное продолжение наружной мембраны клетки. Внутри жгутика строго закономерно располагаются одиннадцать фибрилл. Вдоль оси жгута проходят две центральные фибриллы (рис. 20), берущие начало от аксиальной гранулы. Диаметр каждой из них составляет около 25 нм, а центры их расположены на расстоянии 30 нм. По периферии под оболочкой находятся еще 9 фибрилл, причем каждая слагается из двух тесно спаянных трубочек. Многочисленными исследованиями установлено, что локомоторная деятельность жгутика определяется периферическими фибриллами, тогда как центральные играют опорную функцию и может быть представляют собой субстрат, по которому распространяются волны возбуждения, вызывающие движение жгутика.
|
|
|
Базальное тело, или кинетосома, находится в эктоплазме. Она имеет вид цилиндрического тельца, окруженного мембраной, под которой по периферии кинетосомы располагаются 9 фибрилл, являющихся непосредственным продолжением периферических фибрилл самого жгута. Здесь они, правда, становятся тройными (рис. 20, табл. II, 1).
Иногда основание жгутика продолжается в глубину цитоплазмы за пределы кинетосомы, образуя корневую нить (ризопласт), которая может либо свободно заканчиваться в цитоплазме, либо прикрепляться к оболочке ядра.
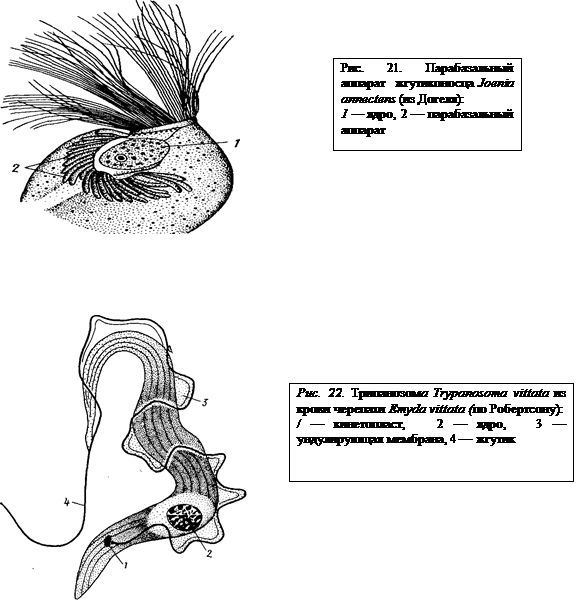
У некоторых жгутиконосцев вблизи кинетосомы располагается парабазальное тело. Форма его может быть разнообразной. Иногда это яйцевидное или колбасовидное образование, иногда же оно приобретает довольно сложную конфигурацию и состоит из множества отдельных долек (рис. 21). Современными электронно-микроскопическими исследованиями доказано, что парабазальное тело — это органоид клетки, гомологичный аппарату Гольджи. Вероятно, в нем конденсируются резервные вещества, служащие энергетической базой локомоторной деятельности жгутика.
 |
У жгутиконосцев, относящихся к отряду Kinetoplastida, у основания жгута кроме кинетосомы помещается еще особый органоид — кинетопласт (рис. 22). По ультрамикроскопической структуре кинетопласт соответствует митохондрии, но, кроме того, содержит значительное количество ДНК. Полагают, что он связан с генерацией энергии для движения жгутика.
|
|
|
У многих растительных жгутиконосцев (эвглены, панцирные жгутиконосцы) имеются особые аппараты, служащие для восприятия световых раздражений. Их называют «глазными пятнами» или стигмами (рис. 23). У эвглены, например, глазное пятно представляет собой скопление мелкозернистого красного пигмента, лежащего близ основания жгутика. У некоторых Dinoflagellata стигма достигает большой величины (до 25 мкм в диаметре) и состоит из чашевидного скопления пигмента, во  вдавлении которого лежит чечевицеобразное зерно крахмала, играющее, вероятно, роль светопреломляющей части глазка — хрусталика.
вдавлении которого лежит чечевицеобразное зерно крахмала, играющее, вероятно, роль светопреломляющей части глазка — хрусталика.
Жгутики служат не только для движения, но и способствуют захвату пищи. Движением жгутика в окружающей воде вызывается водоворот, благодаря которому мелкие взвешенные в воде частички (в том числе бактерии и т. п.) увлекаются к основанию жгутика (рис. 24). Здесь у некоторых жгутиконосцев, питающихся твердой пищей, имеется небольшое отверстие в пелликуле — клеточный рот, ведущий в довольно глубокий канал — глотку, вдающийся внутрь тела. Пища попадает в рот и глотку и далее в Эндоплазме образуется пищеварительная вакуоль. У других видов клеточной глотки нет и у основания жгутика имеется участок липкой цитоплазмы, лишенный пелликулы, через него и происходит восприятие пищи. Непереваренные остатки пищи выбрасываются из тела простейшего. Питание твердой пищей называется анимальным. Однако далеко не все жгутиконосцы питаются твердой пищей. Есть немалое число видов, питающихся жидкой органической пищей и усваивающих ее поверхностью тела. При этом основную роль играют процессы пиноцитоза (см. с. 24). Таковы многие Mastigophora, водящиеся в растворах, богатых разлагающимися органическими веществами. Такой способ питания называют сапрофитным. Анимальное и сапрофитное питание представляет собой две формы гетеротрофного типа обмена веществ, при котором источником пищи являются готовые органические вещества.
 |
Обширные группы жгутиконосцев, а именно растительные жгутиконосцы отр. Phytomastigina, способны к фотосинтезу. Зеленый пигмент хлорофилл локализуется внутри особых тел — хроматофоров, имеющих такое же ультрамикроскопическое строение, как и хлоропласты высших зеленых растений. У одних видов хроматофоров в клетке может быть много, и они имеют форму зерен, у других видов хроматофоров 1—2 и они представляют собой большие изрезанные по краю пластинки. Такие жгутиконосцы способны создавать на свету углеводы из углекислого газа и воды, а также усваивать минеральные соли, в том числе включающие азот и фосфор. Обмен веществ, протекающий за счет энергии света, называется аутотрофным или голофитным. Важно отметить, что некоторые аутотрофные жгутиконосцы (например, эвглены) при известных условиях (в темноте и при наличии в среде растворенных органических веществ) переходят к сапрофитному питанию и могут терять при этом зеленую окраску, вызываемую хлорофиллом. Доказано, что у некоторых эвглен сосуществуют оба типа обмена веществ — аутотрофный и гетеротрофный. Они одновременно осуществляют и фотосинтез, и сапрофитное питание. Такой смешанный тип обмена называется миксотрофным. Столь широкую изменчивость типов обмена веществ у жгутиконосцев следует рассматривать как примитивную особенность, присущую организмам, стоящим как бы на границе между растительным и животным миром.
|
|
|
В теле жгутиконосцев откладываются разного рода резервные питательные вещества. Это могут быть капельки жироподобных веществ, разбросанные в цитоплазме, включения полисахарида гликогена, а у окрашенных растительных жгутиконосцев—зерна крахмала или близкого к нему углевода—парамила (у эвглен).
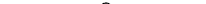 Осморегуляторная и отчасти выделительная функции выполняются у жгутиконосцев, как и у саркодовых (с. 24), сократительными вакуолями, которые имеются у свободноживущих пресноводных форм и отсутствуют у большинства морских и у всех паразитических видов. Клеточное ядро в большинстве случаев присутствует у жгутиконосцев в единственном числе, но существуют также двухъядерные (Lamblia) и многоядерные виды, причем иногда (многие виды Opalina),число ядер может достигать сотни и более.
Осморегуляторная и отчасти выделительная функции выполняются у жгутиконосцев, как и у саркодовых (с. 24), сократительными вакуолями, которые имеются у свободноживущих пресноводных форм и отсутствуют у большинства морских и у всех паразитических видов. Клеточное ядро в большинстве случаев присутствует у жгутиконосцев в единственном числе, но существуют также двухъядерные (Lamblia) и многоядерные виды, причем иногда (многие виды Opalina),число ядер может достигать сотни и более.
|
|
|
Размножение. Для большинства Mastigophora известен только бесполый способ размножения делением надвое.
Деление всегда происходит в продольном направлении, т. е. плоскость деления совпадает с продольной осью тела. Часто деление совершается в свободноподвижном состоянии. Сначала делится митотически ядро, а затем, начиная с переднего конца, постепенно делится все тело простейшего. Что касается жгутикового аппарата, то кинетосома и парабазальный аппарат делятся. Старый жгутик иногда отходит к одной из дочерних особей, а у второй образуется вновь из кинетосомы. В других случаях жгутик отбрасывается и в обеих дочерних особях образуется вновь. Нередко деление совершается в покоящемся состоянии:
простейшее отбрасывает жгутик, округляется и выделяет на своей поверхности прочную оболочку (инцистируется). Под защитой цисты животное последовательно делится один или несколько раз, при этом общий объем тела животного не увеличивается и получаются относительно мелкие клетки.
Процесс последовательных делений без стадий роста и увеличения объема получающихся клеток (процесс напоминает дробление яйца многоклеточных животных) называется палинтомией.Среди жгутиконосцев, в особенности в подклассе Phytomastigina, широко распространена колониальность. Колонии образуются в результате незавершенного деления, когда не вполне отделившиеся друг от друга особи остаются связанными друг с другом. Колонии различаются как по форме, так и по способу развития. На рис. 25 изображена часто встречающаяся в пресных водах древовидная колония Dinobryon. На рис. 26 и 27—разные виды растительных жгутиконосцев, образующих шаровидные колонии. Нередко между особями выделяется слой прозрачного студенистого вещества, и вся колония превращается в полый студенистый шар, в стенке которого в один слой расположены клетки. Число особей, входящих в состав колонии, варьирует от 4 (Gonium) до 10 тыс. и более (Volvox). У такой сложной колонии, как вольвокс, большая часть клеток не дает начала следующему поколению, а периодически умирает, так же как соматические (телесные) клетки многоклеточного организма. Половой процесс имеет место далеко не у всех жгутиконосцев. Он преимущественно распространен у растительных форм, обладающих палинтомическими колониями По характеру развития различают монотомические и палинтомические колонии. Монотомией называют такой способ бесполого размножения простейшего, при котором после акта разделения дочерние особи растут и восстанавливают все органоиды, характерные для материнской клетки. На рис. 26 изображены различные колониальные жгутиконосцы, колонии которых развиваются путем монотомии. Отдельные клетки таких колоний периодически делятся (обычно не все одновременно) и таким образом увеличивается число особей, слагающих колонию. Время от времени сама колония, достигшая предельной для данного вида величины, перешнуровывается пополам. Этим путем осуществляется увеличение числа колоний. У палинтомических колоний, к которым, например, относятся растительные жгутиконосцы семейства вольвоксовых (Volvocidae), размножение происходит иначе. Все клетки колонии Pandorina (рис. 27), Eudorina или только некоторые из них (Volvox) претерпевают последовательные палинтомические деления (см. выше), благодаря чему получается сразу несколько молодых колоний. У Pandorina или Eudorina вся материнская колония при этом распадается на дочерние, число которых равно числу клеток старой колонии. У вольвокса лишь немногие клетки материнской колонии претерпевают палинтомические деления. Образующиеся при этом дочерние колонии сначала помещаются внутри материнской (рис. 28). Через некоторое время старая колония разрушается и расположенные внутри нее молодые становятся свободными. У одиночного бесцветного жгутиконосца Polytoma uvella (рис. 29) копулирующие половые клетки (гаметы) почти не отличимы от вегетативных клеток. Гаметы Polytoma морфологически сходны — различия в строении между мужской и женской гаметами отсутствуют. Такая наиболее примитивная форма полового процесса называется изогамией (равногаметность). Подобная форма полового процесса встречается и у более просто устроенных колоний, например у восьмиклеточной Stephanosphaera. Но у остальных колониальных видов сем. Volvocidae наблюдается постепенный переход к анизогамии (разногаметности), когда между мужскими и женскими гаметами имеются морфологические различия. Уже в 16-клеточной Pandorina одна из копулирующих гамет несколько крупней (женская гамета, или макрогамета) другой, более мелкой (мужская гамета, или микрогамета) (см. рис. 27). У 32-клеточной Eudorina особи одних колоний, целиком не делясь, превращаются в половые—макрогаметы, тогда как особи других дают последовательными делениями (палинтомия) 64 микрогаметы. Копуляция происходит только между мужскими (микрогаметы) и женскими (макрогаметы) половыми клетками. Наибольшее сходство с многоклеточными наблюдается у колоний Volvox (см. рис. 28), у которого не все особи колонии превращаются в гаметы, а лишь немногие. Громадное большинство клеток остается бесполыми. В колонии дают гаметы лишь 25—30 женских клеток и 5—10 мужских. Среди вольвоксов есть раздельнополые виды, у которых на одной колонии развиваются или мужские, или женские гаметы, и гермафродитные, у которых колония дает начало обеим категориям гамет. Каждая из клеток, дающая начало микрогаметам путем палинтомии, производит 256 мельчайших двужгутиковых клеток. Клетки женской ориентации не делятся, но растут и превращаются в крупную макрогамету, которую, как и у многоклеточных организмов, можно назвать яйцевой клеткой. Она неподвижна. Микрогаметы активно отыскивают неподвижные макрогаметы и сливаются с ними. Оплодотворенная макрогамета (зигота) окружается плотной оболочкой. Через некоторое время и при благоприятных внешних условиях она дает начало новой колонии путем последовательных палинтомических делений; старая (материнская) колония разваливается и ее соматические клетки погибают.
 |

У всех жгутиконосцев, имеющих половой процесс, первые два деления зиготы представляют собой мейоз. Таким образом, диплоидна у них лишь зигота, все же остальные стадии жизненного цикла гаплоидны. У жгутиконосцев, следовательно, наблюдается зиготическая редукция в отличие от многоклеточных животных, где мейоз предшествует образованию гамет (гаметическая редукция) и все клетки тела, кроме зрелых половых, диплоидны. В пределах класса жгутиконосцев наблюдаются различные формы полового процесса (копуляция). При этом наблюдаются последовательные этапы от самых примитивных форм изогамии к разным формам анизогамии. У конечного члена этого ряда — вольвокса, происходит формирование настоящих активно подвижных сперматозоидов и яйцевых клеток. Такую форму анизогамии называют согамией.
 Классификация и важнейшие представители. Представители класса жгутиконосцев широко распространены в природе. К нему относятся многочисленные свободноживущие морские и пресноводные, а также паразитические организмы. Ниже мы укажем лишь некоторые наиболее важные отряды этого класса (в современной системе насчитывают 20 отрядов) и несколько отдельных представителей, имеющих особый биологический интерес или практическое значение.
Классификация и важнейшие представители. Представители класса жгутиконосцев широко распространены в природе. К нему относятся многочисленные свободноживущие морские и пресноводные, а также паразитические организмы. Ниже мы укажем лишь некоторые наиболее важные отряды этого класса (в современной системе насчитывают 20 отрядов) и несколько отдельных представителей, имеющих особый биологический интерес или практическое значение.
ПОДКЛАСС I. РАСТИТЕЛЬНЫЕ ЖГУТИКОНОСЦЫ (PHYTOMASTIGINA)
Сюда относятся жгутиконосцы растительной природы, обладающие аутотрофным или миксотрофным способом питания и соответственно несущие зеленый пигмент хлорофилл, с наличием которого связан процесс фотосинтеза. Продуктом ассимиляции чаще всего служит крахмал или близкие к нему полисахариды. В редких случаях хлорофилл может быть утерян и организмы переходят к сапрофитному питанию в богатых растворенными органическими веществами средах.
Отряд 1. Chrysomonadina. Состоит из мелких жгутиконосцев с 1—3 жгутами и с дисковидными хроматофорами золотисто-бурого цвета. Многие из хризомонад способны образовывать псевдоподии. Продукт ассимиляции — полисахарид лейкозин. Обычны в пресных и морских водах. Имеется несколько колониальных видов, из которых в пресной воде часто встречаются древовидные колонии Dinobryon (см. рис. 25) и шаровидные Synura (см. рис. 26).
Отряд 2. Dinoflagellata (Peredinea). Панцирные жгутиконосцы. Двужгутиковые простейшие, обычно обладающие панцирем из клетчатки, слагающимся из закономерно расположенных отдельных пластинок (Peridinium, Ceratium и др.; см. рис. 18). Иногда панцирь отсутствует (рис. 30). Характерно расположение жгутов — оба берут начало рядом друг с другом. Один идет далее назад и свободно выдается в окружающую среду. Другой жгут, называемый поперечным, опоясывает все тело в экваториальной плоскости и располагается в довольно глубоком желобке (см. рис. 18). Большинство панцирных жгутиконосцев снабжено буро-желтыми, реже зелеными, хроматофорами. Тело нередко образует причудливые выросты (см. рис. 18), увеличивающие поверхность и служащие приспособлениями к планктонному образу жизни. Среди Dinoflagellata имеется несколько видов, утерявших хлорофилл и перешедших к гетеротрофному анимальному питанию. Примером могут служить ночесветки (Noctiluca) (см. рис. 30) — обычный компонент морского планктона Черного моря.
 Noctiluca обнаруживает способность светиться при механическом или ином раздражении; процесс связан с окислением жироподобных веществ. Продукт ассимиляции Dinoflagellata— крахмал. Кроме того, в цитоплазме часто имеются в значительных количествах жировые включения. У некоторых морских форм есть глазки и особые защитные стрекательные капсулы с выбрасывающейся стрекательной нитью — яркий пример конвергенции с типом кишечнополостных (с. 117). Панцирные жгутиконосцы в большинстве своем входят в состав морского и пресноводного планктона и играют большую роль в первичной биологической продуктивности Мирового океана. Это определяет их большое значение в биосфере Земли в целом.
Noctiluca обнаруживает способность светиться при механическом или ином раздражении; процесс связан с окислением жироподобных веществ. Продукт ассимиляции Dinoflagellata— крахмал. Кроме того, в цитоплазме часто имеются в значительных количествах жировые включения. У некоторых морских форм есть глазки и особые защитные стрекательные капсулы с выбрасывающейся стрекательной нитью — яркий пример конвергенции с типом кишечнополостных (с. 117). Панцирные жгутиконосцы в большинстве своем входят в состав морского и пресноводного планктона и играют большую роль в первичной биологической продуктивности Мирового океана. Это определяет их большое значение в биосфере Земли в целом.
Отряд 3. Euglenoidea. Содержит большинство наиболее крупных, чаще всего одножгутиковых, окрашенных в зеленый цвет пресноводных жгутиконосцев (см. рис. 23). Некоторые виды рода Euglena в темноте легко обесцвечиваются и переходят от аутотрофного типа обмена к сапрофитному гетеротрофному. Некоторым эвгленам свойствен миксотрофный тип обмена веществ. Продукт ассимиляции — близкий к крахмалу полисахарид парамил.
Отряд 4. Phytomonadina. Включает большое число двужгутиковых зеленых видов с чашевидными хроматофорами. Продукт ассимиляции — крахмал. Преимущественно пресноводные, частью — морские виды. Наряду с одиночными часто очень полиморфными родами (например, род Chlamidomonas включает свыше 300 видов) включает колониальные формы (см. рис. 27, 28).
ПОДКЛАСС II. ЖИВОТНЫЕ ЖГУТИКОНОСЦЫ (ZOOMASTIGINA)
Гетеротрофные жгутиконосцы, ведущие свободноживущий или паразитический образ жизни и обладающие анимальным или сапрофитным способом питания.
Отряд 1. Choanoflagellata. Воротничковые жгутиконосцы. Свободноживущие одиночные (Codosiga, см. рис. 24) или колониальные формы (Sphaeroeca, см. рис. 26, 3, И) с одним жгутиком и расположенным у его основания цитоплазматическим воротничком. Пищевые частицы (бактерии и т. п.) движением жгутика направляются к воротничку, прилипают к нему и постепенно спускаются к его основанию (см. рис. 24). Здесь они переходят в цитоплазму, где в пищеварительных вакуолях подвергаются перевариванию. Электронно-микроскопические исследования показали, что воротничок слагается из множества тесно расположенных друг около друга палочек. Формы колоний воротничковых жгутиконосцев разнообразны: существуют сферические и древовидные колонии.
Отряд 2. Rhizomastigina. Этот небольшой по числу видов отряд свободноживущих простейших интересен тем, что представители его обладают 1—3 жгутами и наряду с этим хорошо развитыми псевдоподиями (см. рис. 17). Они совмещают таким образом признаки двух разных классов.
Отряд 3. Kinetoplastida. Отряд характеризуется наличием особого, связанного со жгутиком органоида—кинетопласта (с. 40). Жгутик чаще всего один, реже их два. Проходя вдоль тела жгутиконосца, жгутик срастается с его поверхностью, образуя при этом ундулирующую мембрану (см. рис. 22). К этому отряду относится небольшое количество свободноживущих видов (виды рода Bodo), но большинство ведет паразитический образ жизни. К ним принадлежит ряд возбудителей опасных болезней человека и животных.
Среди паразитических Kinetoplastida особое внимание заслуживают кровяные паразиты многих позвоночных—трипанозомы (род Truponosoma). Это относительно небольшие жгутиконосцы (см. рис. 22) длиной 20—70 мкм с лентовидным сплющенным телом, заостренным на обоих концах, одним жгутом с ундулирующей мембраной. Трипанозомы живут в крови позвоночных, причем передатчиком их служат различные кровососущие беспозвоночные.
Tryponosoma rhodosiense вызывает в тропической Африке «сонную болезнь» человека. Из западной Африки она постепенно распространилась на восток, на всю экваториальную Африку, погубив за первые три десятилетия XX в. свыше миллиона человек. Начинаясь мало заметной лихорадкой, сонная болезнь постепенно приводит к глубокому истощению и сопровождается сонливостью. При отсутствии лечения она всегда оканчивается смертью. В крови больного человека, лимфатических железах, а позднее и в спинномозговой жидкости можно обнаружить подвижных трипанозом. Природным резервуаром трипанозом служат, по-видимому, антилопы, которые не страдают от присутствия в их организме этих жгутиконосцев. Переносчики трипанозом — кровососущие мухи «цеце» (Glosina morsitans и Gl.palpalis, рис. 31). Муха вместе с кровью больного поглощает трипанозом; последние размножаются в кишечнике мухи и проникают в ее хоботок. Уколом зараженной мухи трипанозомы могут быть переданы в кровь здорового человека.
В настоящее время для лечения трипанозомозов (заболеваний, возбудителями которых являются трипанозомы) успешно используется ряд лекарственных препаратов.
Существует ряд видов трипанозом, вызывающих тяжелые заболевания рогатого скота и верблюдов (Truponosoma evansy, T. brucei). Переносчиками служат кровососущие двукрылые насекомые —слепни и мухи рода Glossina. У лошадей преимущественно на юге (это заболевание констатировано и на территории СССР) наблюдается трипанозомная «случная болезнь». Возбудитель ее— Т. equiperdum передается без переносчика при случке. Болезнь связана с поражением нервной системы.
Родственные трипанозомам лейшмании (Leishmania, рис. 32) представляют собой жгутиконосцев, которые в организме человека являются внутриклеточными паразитами. При этом жгутиковый аппарат их частично редуцирован: паразит имеет вид маленькой округлой клетки (2—4 мкм) с ядром и кинетопластом, но без жгута. Переносчиками служат мелкие кровососущие насекомые — москиты (вид рода Phlebotomis, рис. 33). В кишечнике москита (так же как и при культивировании на
 |
искусственных средах) у паразита появляется жгут. В отличие от трипанозом ундулирующая перепонка отсутствует
Существует два вида лейшманий, патогенных для человека и встречающихся в Средней Азии и Закавказье.

Leishmania donovani вызывает тяжелое заболевание, называемое в Средней Азии «кала-азар» (висцеральный лейшманиоз). Болеют им преимущественно дети. Заболевание сопровождается увеличением печени и селезенки, лихорадкой, малокровием, истощением. Лечение, осуществляемое препаратами сурьмы, обычно успешно.
 |
Другой вид лейшманий — Leishmania tropica вызывает местные заболевания кожи, называемые восточной язвой, или пендинкой. В нашей стране встречается в некоторых районах Закавказья и Средней Азии. После инкубационного периода (от 2 недель до 5 месяцев) на коже (чаще лица и Рис. 33. рук — мест, доступных для укусов москитов) образуется узел, который затем изъязвляется. В ткани язвы под струпом внутри белых кровяных телец находятся многочисленные лейшманий. Продержавшись 1—2 года, язва заживает, оставляя рубец (рис. 34).

В природе имеются естественные источники возбудителей лейшманиозов человека—это собаки, а для кожного лейшманиоза—и грызуны. В норах последних поселяются москиты, передающие паразитов человеку.
Обе формы лейшманиоза (висцеральный и кожный) дают стойкий иммунитет, так что вторичное заболевание невозможно.
Отряд 4. Polymastigina. Исключительно паразитические простейшие, несущие несколько жгутиков, иногда обладающие опорным тяжем — аксостилем и ундулирующей мембраной. В человеке паразитируют виды рода Trichomonas (в кишечнике — T. hominis, в мочеполовых путях — T. vaginalis) (рис. 35). Частым паразитом желчных протоков, двенадцатиперстной и тонкой кишки человека (преимущественно детей) является Lamblia intestinalis (рис. 36). Паразит двусторонне симметричен, имеет два ядра, 8 жгутиков и опорный фибриллярный аппарат, напоминающий аксостиль. Брюшная сторона лямблий образует присоску для присасывания к стенке кишечника. Массовое заражение лямблиями связано с дисфункцией кишечника, иногда с холециститами. Лямблий, попадая в задний отдел
кишечника, сбрасывают жгутики и выделяют толстую оболочку, превращаясь в цисту. В инцистированном состоянии и происходит распространение лямблий. У Trichomonas цисты не найдены.

Отряд 5. Hypermastigina. Сюда относятся многожгутиковые и часто многоядерные формы, обитающие в кишечнике термитов и некоторых тараканов (рис. 37). Большинство Hypermastigina обладает сложной организацией. У них имеются аксостили, часто особые опорные структуры, поддерживающие ядро, сложные парабазальные аппараты.
Биологически Hypermastigina интересны тем, что присутствие их необходимо для нормального пищеварения хозяев. Питаясь почти исключительно древесиной (т. е. клетчаткой), термиты сами по себе не в состоянии ее переваривать. Частицы древесины, попадающие в кишечник термитов, заглатываются и перевариваются многочисленными жгутиконосцами, которые переводят клетчатку в легко усвояемые формы углеводов. Таким образом Hypermastigina — полезные симбионты термитов.
Отряд 6. Opalinina. Крупные, обычно многоядерные (редко двухъядерные) паразитические простейшие, живущие в заднем отделе кишечника различных амфибий. Опалины лишены рта и питаются сапрофитно. Большую роль в этом процессе играет пиноцитоз. Тело опалин покрыто большим количеством (тысячами) жгутиков (ресничек), равномерно распределенных по всей поверхности (рис. 38). Сходство ресничного аппарата опалин с таковым инфузорий (с. 72) долгое время заставляло причислять их к последним в качестве самостоятельного подкласса Protociliata. Однако против принадлежности опалин к инфузориям говорят следующие факты: 1) все ядра у опалин одинаковы, отсутствует характерная для инфузорий дифференцировка их на макронуклеусы и микронуклеусы (с. 75); 2) у опалин, как и у других жгутиковых, половой процесс протекает по типу копуляции. Опалины образуют мелкие одноядерные гаметы. Жизненный цикл опалин поразительно соответствует жизненному циклу хозяев — амфибий. Половой процесс у них происходит раз в году и совпадает с временем размножения хозяина, уходящего для этого в воду. В этот момент в лягушках образуются мелкие цисты опалин, попадающие в воду. Они заглатываются головастиками, в кишечнике которых вышедшие из цист опалины путем ряда делений дают начало одноядерным микро- и макрогаметам (рис. 38). В результате копуляции формируется зигота, в дальнейшем развивающаяся в многоядерную вегетативную форму опалин. Последние размножаются только путем деления. Организация и жизненный цикл опалин настолько своеобразны, что многие ученые выделяют их в самостоятельный подкласс жгутиконосцев, равноценный Phytomastigina и Zoomastigina.
Практическое значение свободноживущих жгутиконосцев. Уже указывалось, что автотрофные жгутиконосцы играют важную роль в биологическом круговороте веществ биосферы как первичные продуценты органического вещества. Это, прежде всего, касается планктонных организмов. Кроме того, многие жгутиковые (как окрашенные, так и бесцветные) служат биологическими индикаторами степени загрязненности (сапробности) вод. По видовому составу жгутиковых (а также других простейших и коловраток) можно составить представление о степени загрязненности водоема органическими веществами. Некоторые виды бесцветных жгутиконосцев, поглощая и усваивая органические вещества (наряду с другими простейшими), играют известную роль в процессе биологической очистки сточных вод.
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 1169; Нарушение авторских прав?; Мы поможем в написании вашей работы!