КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Цитозоль
8-3
8.1.4. Транспорт белков происходит по двум основным путям - через цитозоль и через ЭР [4]
Основные пути транспорта белков показаны на рис. 8-12. Практически все белки образуются на рибосомах, расположенных в цитозоле (кроме нескольких, синтезирующихся на митохондриальных рибосомах хлоропластов). Затем их пути расходятся. Белки, принадлежащие одной транспортной ветви, после завершения их синтеза выделяются в цитозоль. Некоторые из них содержат сигналы сортировки, направляющие их из цитозоля в митохондрии, хлоропласты (у растений), ядро или пероксисомы, другие же - их большинство - не имеют специфических сигналов сортировки и остаются в цитозоле в качестве постоянных компонентов.
Другой основной путь транспорта используется при синтезе белков, предназначенных для выведения из клетки, а также белков, которые должны стать компонентами ЭР, аппарата Гольджи, плазматической мембраны или лизосом. Все эти белки по мере их образования переносятся в ЭР при помощи сигналов сортировки, расположенных обычно на N-конце. Рибосомы, на которых собираются такие белки, остаются связанными с мембраной ЭР в течение недолгого времени после начала синтеза полипептидной цепи. Как только очередной участок полипептидной цепи синтезируется, он проникает через липидный бислой этой мембраны. Некоторые белки затем попадают в просвет ЭР, другие остаются частично заключенными в мембрану в качестве трансмембранных белков.
Обычно проходит не больше одной или двух минут с момента высвобождения белка в цитозоль до поступления его в соответствующую органеллу. Белки, предназначенные для ядра, митохондрий или
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
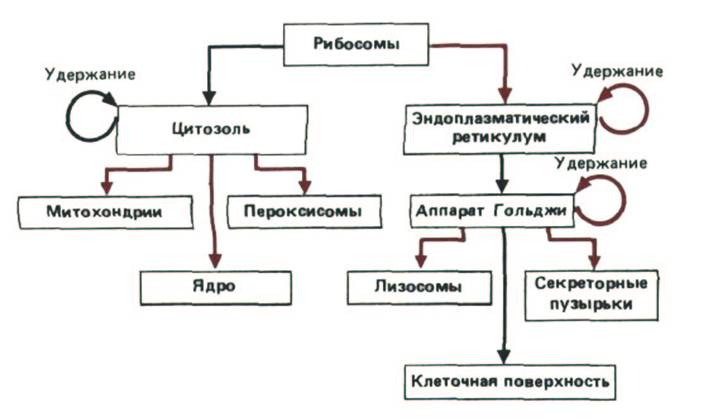
Рис. 8-12. Упрощенная схема путей метаболизма белков. Сигналы, направляющие движение конкретного белка по определенной «дороге» и, следовательно, определяющие локализацию этого белка в клетке, содержатся в его аминокислотной последовательности.
«Путешествие» начинается с синтеза белка на рибосоме и заканчивается его прибытием к месту конечного назначения. На каждой промежуточной стадии (изображены прямоугольниками) принимается решение - оставить данный белок на ней или транспортировать дальше. В принципе, сигнал может требоваться либо для удержания белка в каждом из указанных компартментов, либо для его продвижения дальше, а альтернативное событие может происходить «по умолчанию» (т. е. без специального сигнала). На этой схеме те пути, которые, вероятнее всего, требуют специальных сигналов, выделены цветом; те же пути, которые, скорее всего, выбираются по умолчанию, показаны черными стрелками. Так как образование эндосом изучено плохо, здесь не изображено никаких путей, ведущих к эндосомам.
пероксисом, достигнув этих органелл, на этом заканчивают свой путь, между тем белки, попадающие в ЭР, вовлекаются в дальнейший транспорт, который, как полагают, происходит с помощью транспортных пузырьков, отделяющихся от одной мембраны и сливающихся с другой. По-
видимому, для достижения конечного пункта должно произойти несколько таких циклов, и путь от мембраны ЭР до места назначения может занять один час.
8.1.5. Белки могут перемешаться между компартментами двумя принципиально различными способами [5]
Чтобы понять общие принципы работы сигналов сортировки, важно различать два совершенно различных пути, по которым белки перемещаются из одного компартмента в другой. Во-первых, они могут непосредственно проникать через мембрану, попадая из пространства, топологически эквивалентного цитозолю, в пространство, топологически эквивалентное внеклеточному, или наоборот. Этот путь требует наличия в мембране специального белка-транслокатора, кроме того, молекула транспортируемого белка должна развернуться, чтобы, подобно змее, «проползти» сквозь мембрану. В качестве примера такого рода событий может служить перемещение определенных белков из цитозоля в просвет ЭР. Второй путь передвижения белковых молекул опосредован транспортными пузырьками. Эти пузырьки захватывают определенные молекулы в полости одного компартмента (от которого они отшнуровываются) и переносят их в другой компартмент, сливаясь с ним. Именно так происходит перенос растворимых белков из ЭР к аппарату Гольджи (см. рис. 8-Ю). При таком везикулярном транспорте белки не пересекают никаких мембран, поэтому они переносятся только между компартментами, топологически эквивалентными друг другу.
Оба типа транспортных процессов избирательно контролируются с помощью специальных белков, выполняющих роль сигналов сортировки. У белка, который напрямую переносится через мембрану, эти сигналы распознаются транслокатором в мембране. А в транспортный пузырек белок попадает, если его сигнал сортировки связывается с рецептором на мембране пузырька. Вероятно, существуют и такие транспортные пузырьки, которые способны захватывать белки, утерявшие специфические сигналы сортировки. В любом случае вновь образующиеся пузырьки переносят только предназначенные для этого белки.
В настоящее время некоторые сигналы сортировки в составе белков известны, тогда как большинство комплементарных им мембранных рецепторов - нет. Кроме того, мы не знаем почти ничего о транспортных
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
пузырьках. Можно предположить только, что их существует множество видов, в соответствии с различными переносами, которые они должны осуществлять. Чтобы выполнять свои функции, каждый пузырек должен захватывать только определенные белки и сливаться только с определенной мембраной-мишенью: например, пузырек, переносящий молекулы от ЭР к аппарату Гольджи, не должен включать белки, которые останутся в ЭР, и должен сливаться только с аппаратом Гольджи, а не с другими органеллами. В этой главе мы рассмотрим, каким образом транспортные пузырьки достигают такой избирательности.
8.1.6. Сигнальные пептиды и сигнальные участки определяют судьбу белка [6]
Полагают, что на белках существуют два типа сигналов сортировки, направляющие их, шаг за шагом, вдоль разветвляющихся путей (рис.
8-12). Для некоторых стадий сигналы сортировки представляют собой протяженный участок аминокислотной последовательности длиной 15-60
остатков. Когда эта стадия пройдена, такой сигнальный пептид отрезается. Сигналом сортировки для других стадий, вероятно, служит определенная трехмерная структура, образуемая атомами поверхности белка при свертывании его молекулы. Аминокислотные остатки, формирующие такие сигнальные участки, могут быть расположены очень далеко друг от друга в линейной последовательности белка (рис. 8-13).
Сигнальные пептиды направляют белки из цитозоля в ЭР, митохондрии, хлоропласты и ядро; они также отвечают за то, чтобы некоторые белки остались в ЭР. Сигнальные участки, видимо, играют важную роль при распознавании определенных лизосомных белков специальным ферментом в аппарате Гольджи.
Чтобы выяснить пункт назначения того или иного белка внутри клетки, необходимо определить тип его сигнального пептида (табл. 8-3).
Белки, которые должны попасть в ЭР, обычно несут N-концевой сигнальный пептид. Его центральная часть образована 5-Ю гидрофобными аминокислотными остатками. Большинство этих белков направляется из ЭР в аппарат Гольджи; те же, которые имеют на С-конце специфическую последовательность из четырех аминокислот, остаются в качестве постоянных компонентов. Многие белки, предназначенные для митохондрий, имеют сигнальные пептиды, в которых положительно заряженные аминокислотные остатки чередуются с гидрофобными. Среди белков, направляющихся в ядро, большинство имеет сигнальные пептиды, образованные кластером положительно заряженных аминокислотных остатков.
Наконец, некоторым белкам цитозоля присущи сигнальные пептиды, с которыми ковалентно связывается жирная кис-
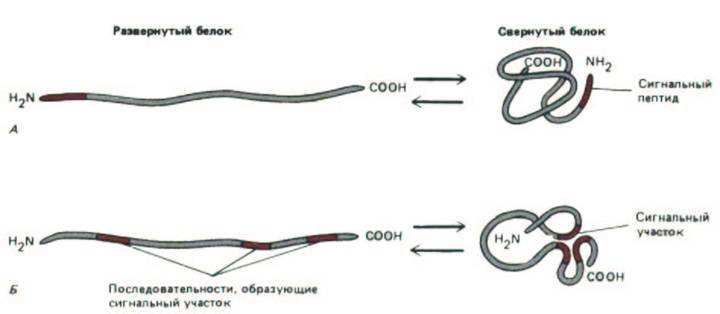
Рис. 8-13. Два способа образования на белке транспортного сигнала. А. Сигнал представляет собой единственный дискретный отрезок аминокислотной последовательности, называемый сигнальным пептидом. В свернутом белке сигнальный пептид располагается снаружи.
Сигнальные пептиды чаще находятся на конце полипептидной цепи (как показано здесь), но могут быть и в других местах. Сигнальные пептиды выявляют экспериментально по их эффекту на внутриклеточную сортировку белков. Для этого сигнальный пептид с помощью методов рекомбинантных ДНК «пришивают» к какому-нибудь другому белку. Б. Сигнальный участок может быть сформирован при наложении аминокислот из удаленных друг от друга районов при сворачивании молекулы; или же такой сигнал образуется из отдельных «пятен» на поверхности свернутого белка, удаленных друг от друга на известное расстояние. И в том, и в другом случае транспортный сигнал зависит от трехмерной конформации белка. По этой причине точно локализовать такие сигналы чрезвычайно трудно.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
Таблица 8-3. Типичные последовательности сигнальных пептидов
| Функция сигнального пептида | Пример сигнального пептида |
| Импорт в ЭР | 
|
| Оставление в просвете ЭР | 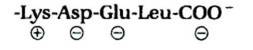
|
| Импорт в митохондрии | 
|
| Импорт в ядро | 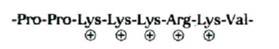
|
| Присоединение к мембране путем ковалентного связывания миристиновой кислоты с N-концом | 
|
Заряженные остатки обозначены + или —. Большой блок гидрофобных остатков заключен в рамку. H3N- обозначает N-конец белка; —
СOO- обозначает карбоксильный конец лота, направляющая эти белки к мембранам без проникновения в ЭР.
Роль каждого такого сигнального пептида в доставке белка по месту назначения была продемонстрирована в опытах по генной инженерии. Например, помещение N-концевого сигнального пептида, связывающегося с ЭР, в начало белка цитозоля, заставляло этот белок направляться к ЭР. Сигнальные пептиды белков, имеющих одно и то же конечное назначение, функционально взаимозаменяемы, хотя их аминокислотные последовательности могут сильно различаться. Возможно, для процесса распознавания сигнала физические свойства (такие, например, как гидрофобность) оказываются важнее, чем точная последовательность аминокислот.
Сигнальные участки анализировать гораздо труднее, чем сигнальные пептиды, поэтому об их структуре известно гораздо меньше.
Поскольку они образуются из сложной трехмерной конфигурации свернутого белка, их невозможно просто перенести от одного белка к другому.
Более того, экспериментальное повреждение сигнального участка часто влечет за собой повреждение белка как целого.
8.1.7. Клетки не могут строить свои мембранные органеллы de novo: им требуется информация, содержащаяся в
самой органелле [7]
Когда клетка воспроизводится и делится, она должна удваивать свои мембранные органеллы. Обычно это происходит путем увеличения размеров этих органелл при включении в них новых молекул; увеличенные органеллы затем делятся и распределяются по двум дочерним клеткам.
Вряд ли клетка в состоянии создавать эти органеллы de novo. Если, например, из нее полностью удалить ЭР, она не сумеет его реконструировать.
Ведь мембранные белки, определяющие специфику ЭР и выполняющие множество его ключевых функций, сами являются продуктами ЭР: без ЭР или по крайней мере без мембраны, содержащей
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
транслокаторы, необходимые для импорта белков в ЭР (и при отсутствии транслокаторов, которые требуются для импорта белков в другие органеллы), новый ЭР не может быть построен.
Следовательно, для формирования мембранных органелл недостаточно только информации ДНК, определяющей белки органелл.
Необходима также «эпигенетическая» информация в виде хотя бы одного характерного белка в мембране органеллы. Эта информация передается от родительской клетки потомству с самой органеллой. Вероятно, такая информация необходима для поддержания компартментации клетки, тогда как информация, содержащаяся в ДНК, необходима для «размножения» нуклеотидных и аминокислотных последовательностей.
Заключение
Эукариотические клетки содержат внутриклеточные мембраны, замыкающие около половины общего объема клетки в отдельные внутриклеточные компартменты. Основные виды мембранных органелл во всех эукариотических клетках следующие: эндоплазматический ретикулум, аппарат Гольджи, ядро, митохондрии, лизосомы, эндосомы и пероксисомы; растительные клетки содержат также хлоропласты. Каждая органелла имеет в своем составе различные белки, определяющие ее уникальные функции.
Каждый вновь синтезированный белок органелл проходит от рибосомы до органеллы особый путь, определяемый либо сигнальным пептидом, либо сигнальным участком. Сортировка белков начинается с первичной сегрегации, при которой белок либо остается в цитозоле, либо переносится в другой компартмент (например, в ядро, митохондрию или эндоплазматический ретикулум). Белки, попадающие в ЭР, претерпевают дальнейшую сортировку по мере того, как они переносятся в аппарат Гольджи и затем из аппарата Голъджи в лизосомы, в секреторные пузырьки или к плазматической мембране. Некоторые белки остаются в ЭР и в различных цистернах аппарата Гольджи. Белки, предназначенные для других компартментов, видимо, попадают в транспортные пузырьки, которые отшнуровываются от одного компартмента и сливаются с другим.
Цитозоль - это часть цитоплазмы, занимающая пространство между мембранными органеллами. Обычно на него приходится около половины общего объема клетки (см. табл. 8-1). В состав цитозоля входит множество ферментов промежуточного обмена, кроме того, он заполнен рибосомами, синтезирующими белки. Около половины всех белков, образующихся на рибосомах, остаются в цитозоле в качестве его постоянных компонентов. В данном разделе мы обсудим судьбу этих белков, а также некоторые механизмы, позволяющие контролировать их время жизни и направлять белки в определенные места цитозоля.
8.2.1. Организация цитозоля поддерживается белковыми филамептами [8]
Цитозоль содержит множество белковых филаментов, собранных в фибриллярный цитоскелет (гл. 11). Именно цитоскелет определяет форму клетки, обеспечивает движение цитоплазмы и образует общую сеть, которая организует ферментативные реакции. Более того, поскольку белки составляют около 20% массы цитозоля, правильнее будет представлять его себе как высокоорганизованный гель, а не как pacтвор
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
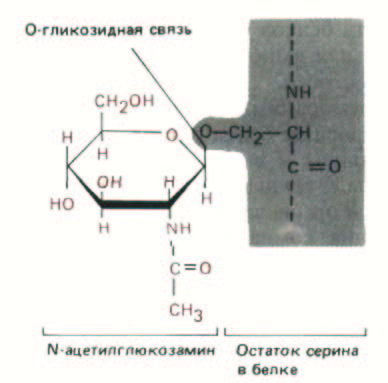
Рис. 8-14. Единственной известной формой гликозилирования, протекающего в цитозоле клеток млекопитающих, является присоединение N-ацетилглюкозамина к остаткам серина или треонина в белке. Таким способом модифицируются белки-регуляторы многих генов и некоторые белки ядерных пор; назначение этой модификации неизвестно. В ЭР и аппарате Гольджи происходит гораздо более сложное гликозилирование (см. рис. 8-52).
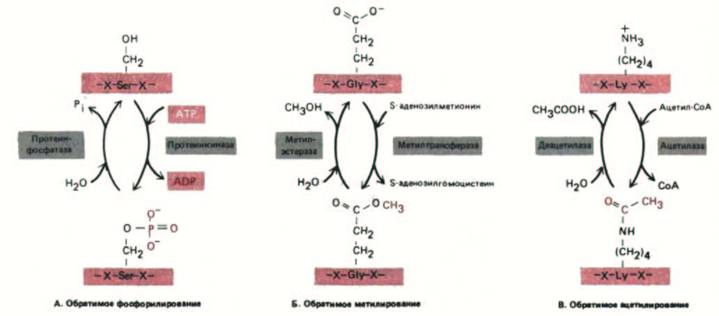
Рис. 8-15. Три типа обратимых ковалентных модификаций, которым подвергаются белки и которые влияют на их активность. Каждая изображенная модификация изменяет заряд боковой цепи аминокислоты. Наиболее распространенная модификация - это фосфорилирование ОН-
групп боковых цепей серина, треонина и тирозина в белке. По существующим оценкам, в животных клетках этим способом модифицировано около 10% белков цитозоля.
ферментов. Исследования скорости диффузии, однако, показывают, что малые молекулы и некоторые небольшие белки диффундируют в цитозоле почти с той же скоростью, что и в дистиллированной воде. Таким образом, с точки зрения промежуточного обмена (где и субстраты, и продукты являются малыми молекулами) мы можем рассматривать цитозоль как простой раствор.
С другой стороны, известно, что большие частицы, такие, как транспортные пузырьки и органеллы, движутся очень медленно, отчасти потому, что часто сталкиваются с компонентами цитоскелета. Чтобы они передвигались с приемлемой скоростью, специальные белковые «моторы»
гидролизуют АТР и используют освобождающуюся при этом энергию для переноса крупных частиц вдоль микротрубочек или актиновых филаментов. Специфические филаменты выполняют в данном случае роль «рельсов», направляющих транспортные пузырьки к соответствующей мембране-мишени. Большинство исследователей клетки, однако, считают функцию цитоскелета менее специализированной и полагают, что специфичность везикулярного транспорта обеспечивается в основном системами рецепторов, расположенных на обращенной к цитозолю поверхности самих пузырьков (см. разд. 8.8.6).
8.2.2. Многие белки претерпевают в цитозоле ковалентные модификации [9]
Описано более 100 различных посттрансляционных модификаций белков. Роль большинства этих модификаций не выяснена; некоторые из них случайны и, по-видимому, не имеют функционального значения, но есть и такие, которые важны для жизни клетки, так как они тщательно контролируются специфическими ферментами. Далее мы увидим, что некоторые модификации происходят в ЭР и аппарате Гольджи. В этих органеллах, например, ферменты гликозилирования добавляют к белкам сложные цепи остатков Сахаров, образуя гликопротеины (см. разд. 8.6.12).
Единственный известный случай гликозилирования в цитозоле клеток млекопитающих - это добавление к белкам N-ацетилглюкозамина (рис. 8-14).
Однако множество других ковалентных модификаций протекает в первую очередь именно в цитозоле. Некоторые из них стабильны, и необходимы для активности белка, например, ковалентное
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
присоединение коферментов (биотина, липоевой кислоты или пиридоксальфосфата). Определенные ковалентные модификации, происходящие в цитозоле, обратимы и служат для регуляции активности многих белков (рис. 8-15).
Среди известных в настоящее время модификаций описана одна, чрезвычайно важная для доставки белков к месту назначения.
Присоединение жирной кислоты к белку направляет его к определенным мембранам, обращенным в цитозоль.
|
|
Дата добавления: 2015-08-31; Просмотров: 1762; Нарушение авторских прав?; Мы поможем в написании вашей работы!